论文信息
英文题目:Reversible kink instability drives ultrafast jumping in nematodes and soft robots
中文题目: 可逆扭结不稳定性驱动线虫与软体机器人的超高速跳跃
作者:Sunny Kumar, Ishant Tiwari, Victor M. Ortega-Jimenez, Adler R. Dillman, Dongjing He, Yuhang Hu, Saad Bhamla(通讯)
作者单位: 佐治亚理工学院、加州大学河滨分校、加州大学洛杉矶分校
期刊:Science Robotics(IF 27.5中科院一区,JCR Q1)
发表时间:2025年4月23日
链接:https://www.science.org/doi/10.1126/scirobotics.adq3121
引文格式:Kumar S, Tiwari I, Ortega-Jimenez V M, et al. Reversible kink instability drives ultrafast jumping in nematodes and soft robots[J]. Science Robotics, 2025, 10: eadq3121.
01 全文速览
一根吸管弯到一定程度会“打结”,这在工程上通常意味着失效。但有一种比头发丝还细的线虫,偏偏靠这种“打结”来完成超高速跳跃——并且跳出了20倍体长的高度。
佐治亚理工学院的团队发现,昆虫病原线虫(Steinernema carpocapsae)利用一种可逆的扭结不稳定性来突破肌肉力量的极限。它们先用1.5秒慢速形成一个α形环,然后在70微秒内突然弹开,起跳速度达到1.5米/秒,输出功率高达10⁴瓦/公斤——是同样大小的秀丽隐杆线虫的10万倍。
更关键的是,他们把这套机制“翻译”成了工程语言。通过仿生模型(SoftJM),他们验证了三个核心设计原则:可逆扭结让系统能在有限力量下储存更多弹性能;液体门控利用表面张力充当“锁扣”,实现微秒级释放;长径比调节让线虫主动改变身体几何,触发扭结并提升跳跃效率。
基于这些发现,他们设计出含碳纤维骨架的软体跳跃机器人SoftJM 4,实现了25倍体长的跳跃高度,并展示了沙地跳跃、磁吸壁挂、安全碰撞等能力。这项研究揭示了一种全新的软体机器人驱动范式——把通常被视为失效模式的“扭结”,变成了高效能量释放的开关。
核心亮点:
✅可逆扭结:线虫主动形成扭结,让弯曲力在达到上限后不再增加,却能继续储存能量——这种“力饱和”机制让有限肌肉力发挥最大储能效果
✅液体门控:微米级水膜充当“锁扣”,70微秒内破裂释放能量,时间尺度比弹指快50倍
✅主动调长径比:线虫把身体长径比从20主动压缩到7以下,触发扭结并提升跳跃效率
✅刚柔并济:碳纤维骨架+硅胶软体,既满足储能刚度和功率输出,又保持与人交互的安全性
✅环境适应:在沙子等粗糙表面也能稳定起跳,展示了软体机器人野外应用的潜力
02 研究内容
🧬扭结:从失效模式到驱动开关
提起“扭结”,你可能想到的是弯过头的水管、吸管,或者折弯的塑料片——这些通常意味着结构失效。但线虫偏偏把它变成了一个可逆的储能开关。
图1展示了线虫跳跃的全过程。A图是线虫静息状态(体长0.5毫米,直径20微米,长径比25:1)。B图是关键的“扭结”状态:线虫把身体弯成α形环,在腹侧形成高曲率扭结。C图是弹开后腾空,高度超过20倍体长。
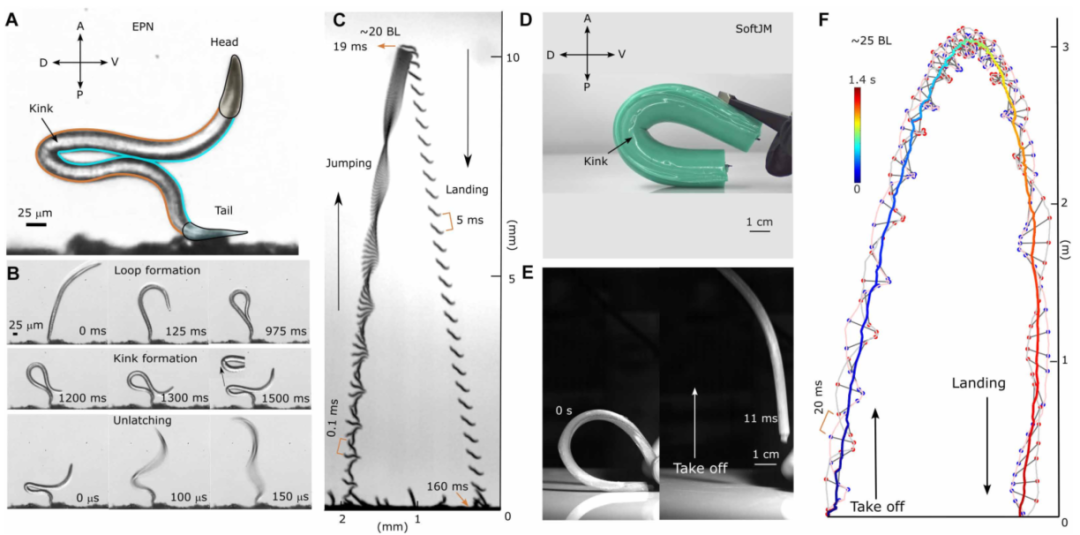
图1 跳跃线虫和仿生软跳跃模型。 (A) EPN S. carpocapsae 处于弯曲或“扭结”位置的图像,是在跳跃之前通过高速摄像机的放大视图拍摄的。比例尺为 25 μm。 P,后部; D,背部; A,前部; V,腹侧。 (B) 顺序图像显示了 EPN 的跳跃准备:从 0 到 975 ms 的环路形成,从 1200 ms 到 1500 ms 的扭结形成,以及持续 150 μs 的最终解锁阶段。比例尺为 25 μm。 (C) 以 10,000 fps 记录的起飞(速度~1.5 m/s)、旋转和着陆(速度~0.15 m/s)期间 EPN 的轨迹。 (D) SoftJM 代表在宏观尺度上复制 EPN 跳跃的物理模型。 (E) 显示 SoftJM 起飞阶段的图像。 (F) SoftJM 的轨迹以 240 fps 捕获,展示了起飞、旋转和着陆的不同阶段。
这个扭结不是线虫被动“弯过头”的结果,而是主动调控的。它们用1.5秒缓慢收紧腹-腹接触的α环,然后在70微秒内突然弹开。起跳速度1.5米/秒,功率密度10⁴瓦/公斤——对比一下,秀丽隐杆线虫的肌肉功率密度只有0.1瓦/公斤量级。这5个数量级的差距,暗示线虫用了某种弹簧-门控机制,而不是纯肌肉驱动。
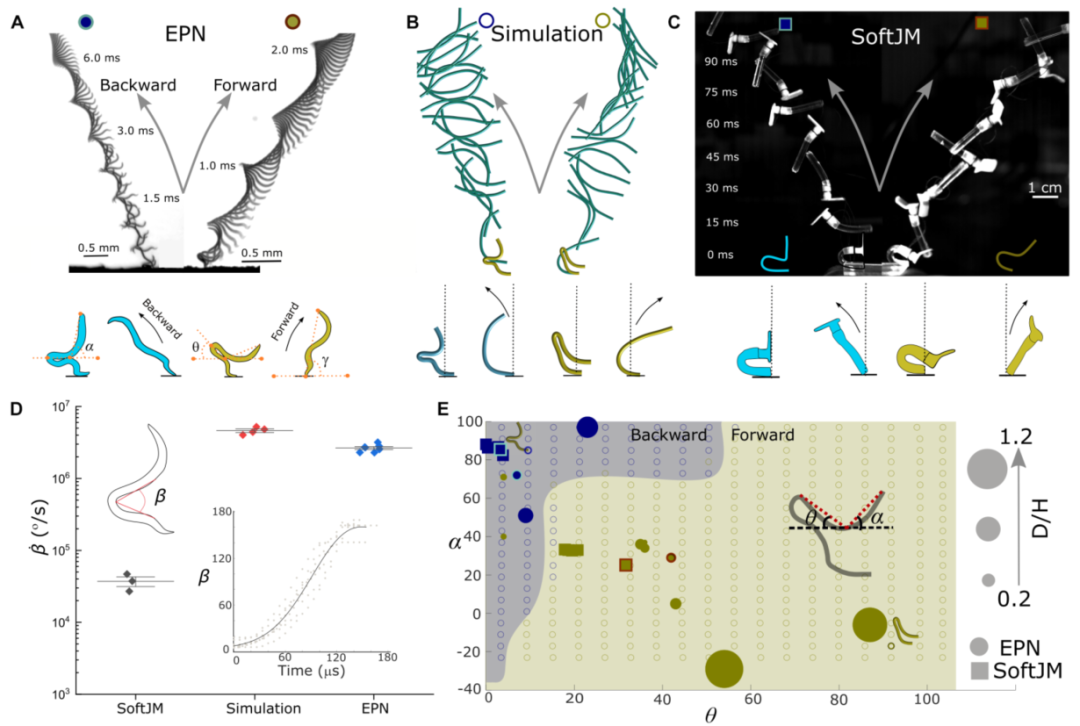
图2 EPN的定向跳跃,SoftJM和模拟。(A)EPN向后和向前跳跃。EPN的起飞角度(γ)取决于其姿态,由角度θ和α量化。调整这些角度可以相对于其与基板的接触点向后或向前移动其COM,从而导致向后跳跃(α=∞90°,θ=∞0°,γ=∞120°)或向前跳跃(α=∞45°,θ=∞45°,γ=∞60°),更多详情请参见图S3A和表S5。(B和C) Cosserat杆模拟和软跳高运动员的物理模型说明了向前和向后方向的跳跃。(d)soft JM(n = 3个人)、模拟(n = 4次实现)和EPNs (n = 7个人)的角速度(̇ β)。插图显示了作为EPN(n = 5)的时间函数绘制的张角β。(E)模拟EPN不同姿态的模拟结果,用不同颜色表示向后(蓝环)和向前(黄环)起飞。圆圈代表EPN跳跃,而正方形代表SoftJM跳跃。这些纯色数据点的大小与其归一化轨迹范围(D/H)成比例。
🔬扭结如何“锁”住能量?
是什么“锁”住了这个扭结?又是怎么在微秒内解开的?
答案出乎意料:水膜。早在1965年,Reed和Wallace就猜测线虫腹-腹接触点可能有水滴起固定作用。50多年后,高速摄像证实了这一点。
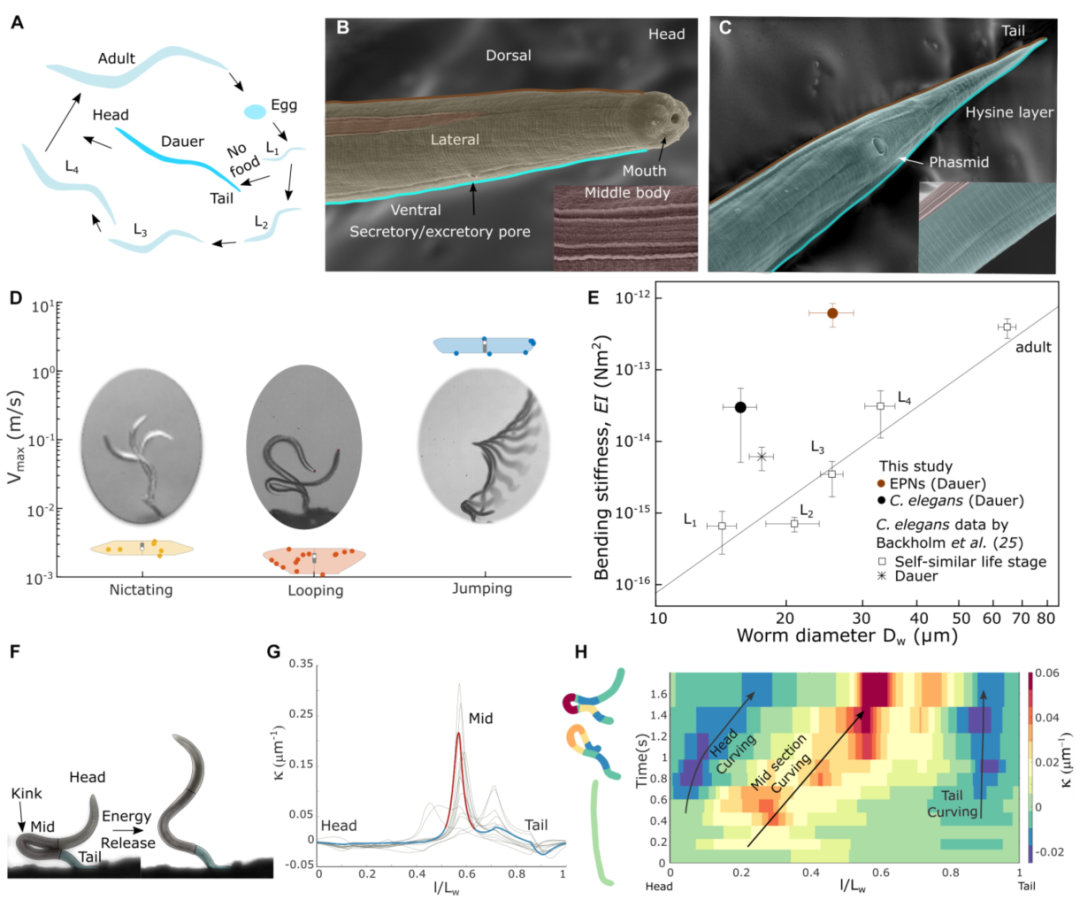
图3 线虫的弯曲刚度和曲率分析。(A)线虫的生命阶段,从幼虫阶段L1到L4和成虫阶段,而另一个非摄食幼虫阶段(道伊尔)生命周期是从L1到道伊尔再到成虫阶段。(B)一只EPN的扫描电子显微镜(SEM ),突出显示了一只受感染幼体(dauer)的头部和颈部。黑色箭头表示头部的嘴和分泌孔。插图显示了EPN车身的侧面。(C)EPN尾部的扫描电镜,插图显示线虫的腹侧。(D)小提琴图显示了在瞬动(n = 6)、成圈(n = 14)和跳跃(n = 6)期间,成圈和瞬动之间(P < 0.01)以及成圈和跳跃之间或瞬动和跳跃之间(P < 0.0001)的EPN的最大速度分布。(dauer阶段EPN和C. elegans的弯曲刚度,通过AFM测量,作为线虫直径𝐷w的函数(n = 3个个体,每个个体三次)。比较包括Backholm等人(25)报道的秀丽隐杆线虫每个生命阶段的平均自相似线虫。与秀丽隐杆线虫的相同自相似生命阶段和涂抹子相比,涂抹子阶段EPN表现出更高的刚度。(F)在跳跃和打开环之前EPN的不同区域(头部、中部和尾部)。(g)在跳跃(n = 14)之前,具有标准化长度(l/Lw)的EPN的最终状态曲率,其中𝑙是穿过线虫体的变化长度,Lw是EPN的总长度。(H)基模图显示了作为时间函数的EPN长度上的曲率增长。左侧的彩色编码EPN示意图显示了线虫身体的弯曲轮廓。
图4A-D展示了水膜从形成到破裂的全过程。D图显示,水膜在约70微秒内迅速变薄、破裂,扭结瞬间弹开。这个时间尺度比弹指(约7毫秒)快了两个数量级。
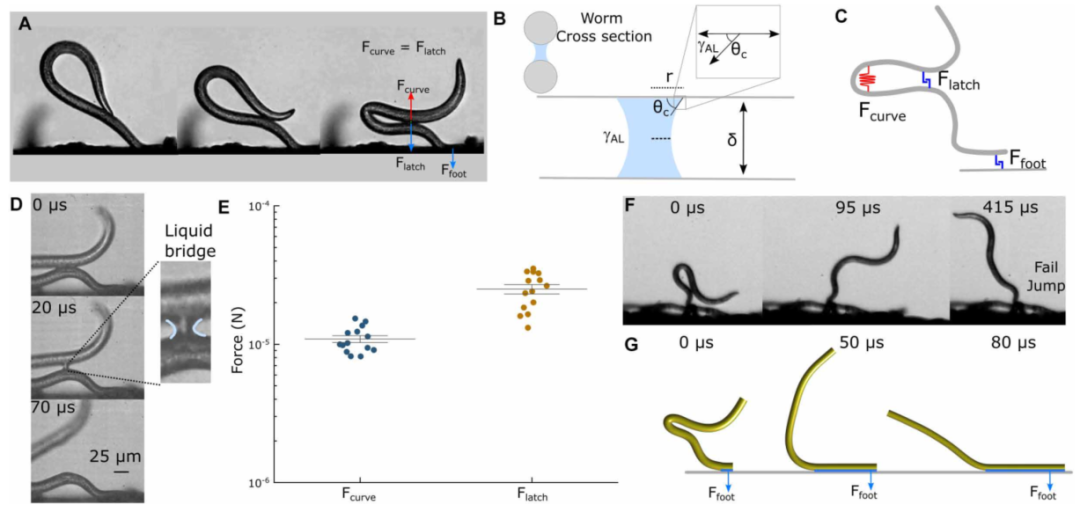
图4 液体闩锁机构和力平衡。(A) EPN在腹侧-腹侧接触处形成液体栓。(B)假设完全润湿(θc = 0°),液体表面张力用γAL表示的液桥横截面图像。(c)一个简化的示意图,显示了参与EPN跳跃的力:毛细管闩锁𝐹latch、曲率力𝐹curve和由于毛细管力𝐹Foot.引起的尾部粘附(D)在EPN的腹腹接触期间,随着时间的推移,液桥变薄和断裂。(e)散点图表示由于曲率产生的弯曲力𝐹curve和由于腹侧-腹侧接触之间的液体桥产生的毛细力𝐹latch(n = 15)。(F)由于尾部附着在表面上,跳跃失败。(G)模拟快照,显示模拟尾翼附着导致的失败跳跃。
他们建立了一个力平衡模型:弯曲力
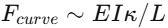
,毛细力
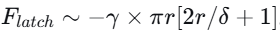
。计算发现,这两种力都在10⁻⁵牛量级——水膜提供的毛细力正好能“锁住”扭结。当线虫持续收紧时,弯曲力超过毛细力阈值,水膜破裂,能量释放。
还有一个细节:如果尾部的液体桥黏附力过大,线虫即使打开了腹侧水膜,也可能跳不起来(图4F-G)。这解释了为什么环境湿度太高时,线虫跳不成功——不是扭结没形成,而是尾部被“粘”住了。
🧪仿生验证:为什么扭结反而能存更多能?
为了验证扭结的力学价值,他们设计了一组仿生模型SoftJM(图5A)。
图5B-D是弯曲测试。SoftJM 2(水球+塑料条)的恢复力-压缩比曲线有一个转折点:扭结形成前,力随弯曲线性增长;扭结形成后,力增长变缓,几乎进入“力饱和”平台期。这意味着线虫在肌肉力有限的情况下,可以继续往身体里“充”弹性能,而不需要肌肉力继续增大。
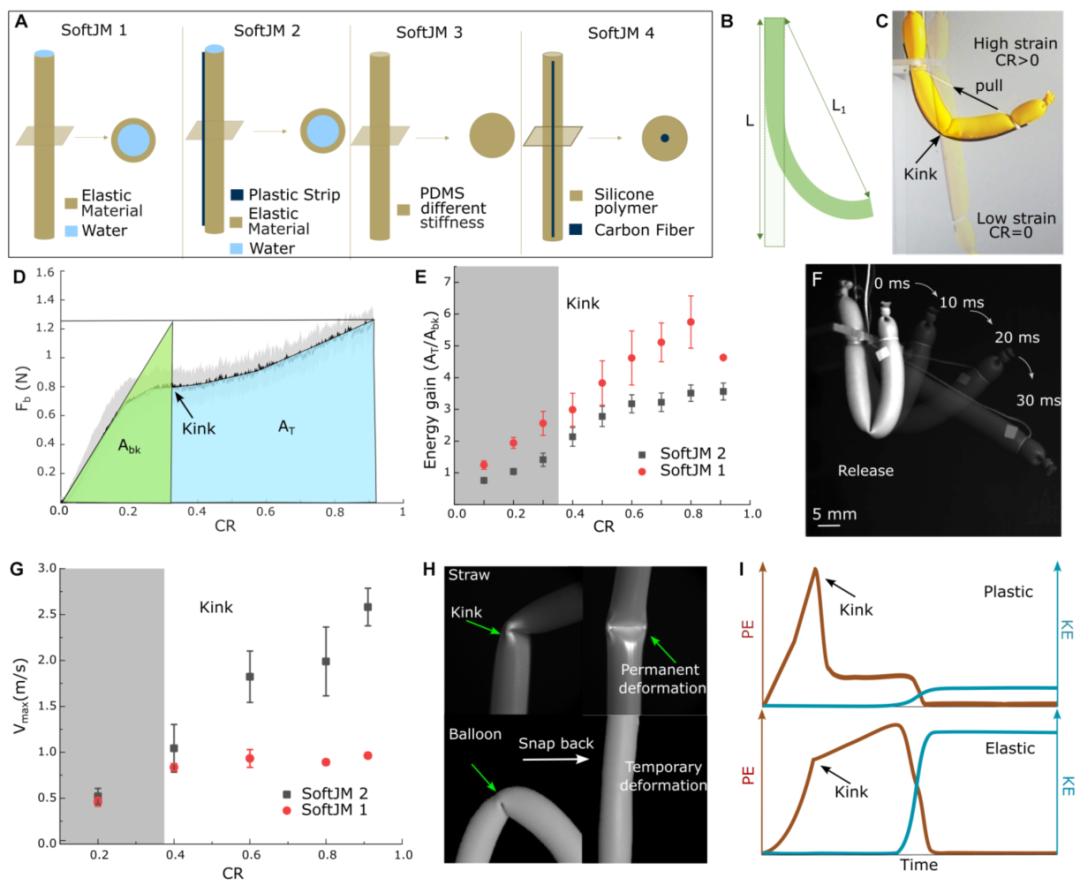
图5 软JMs中扭结诱导的弹性能量存储和力动力学。(A)各种合成软跳跃模型(SoftJM): SoftJM 1(水球,低刚度),SoftJM 2(水球+条带,刚性骨架),SoftJM 3 (PDMS,不同的杨氏模量,见表S3),和SoftJM 4(硅酮聚合物+ CF)。(b)测量弯曲力的杆弯曲示意图(𝐹b),其中𝐶𝑅=(𝐿-𝐿1)/𝐿.(C)在弯曲长宽比η = 10的SoftJM 2时形成扭结。(d)soft JM 2在不同CR下的𝐹b阴影误差曲线(黑线,η = 10,n = 3),其中黑色实线代表平均力响应,周围灰色阴影区域代表实验SD。(e)soft JM 1和SoftJM 2的能量增益(𝐴T/𝐴bk)散点图(n = 5,η = 10)。(F)soft JM 1在不同时间间隔释放后的可视化,η = 10。(G) 𝑉max散点图(y轴SD)显示了不同物理模型SoftJM 1(红点)和SoftJM 2(黑点)的最大骤回速度(n = 3,η = 10,P < 0.0001)。(H)如果出现暂时变形(弹性气球或PDMS),圆柱形杆会弯曲并迅速弹回;否则,它会发生永久变形(塑料吸管)。(I)代表输入势能(PE)的能量平衡示意图;红线)和输出动能(KE;蓝线)表示不可逆(塑性)和可逆(弹性)系统。
图5E显示,SoftJM 1(水球)和SoftJM 2(水球+塑料条)相比无扭结的理想线性弹簧,实际储存的能量分别提升了5倍和4倍。更重要的是,图5G显示扭结后弹开的最大速度随压缩比增加而提升,说明扭结不是“耗能”的失效,而是让能量释放更快。
🎯主动调长径比:一个被忽视的控制参数
线虫不仅能“打结”,还能主动调节身体几何。图6A-B展示了这个过程:初始长径比约20,形成α环后有效长径比降到7以下。图6G显示,长径比越小,最大弯曲力越大——这意味着储能容量越大。
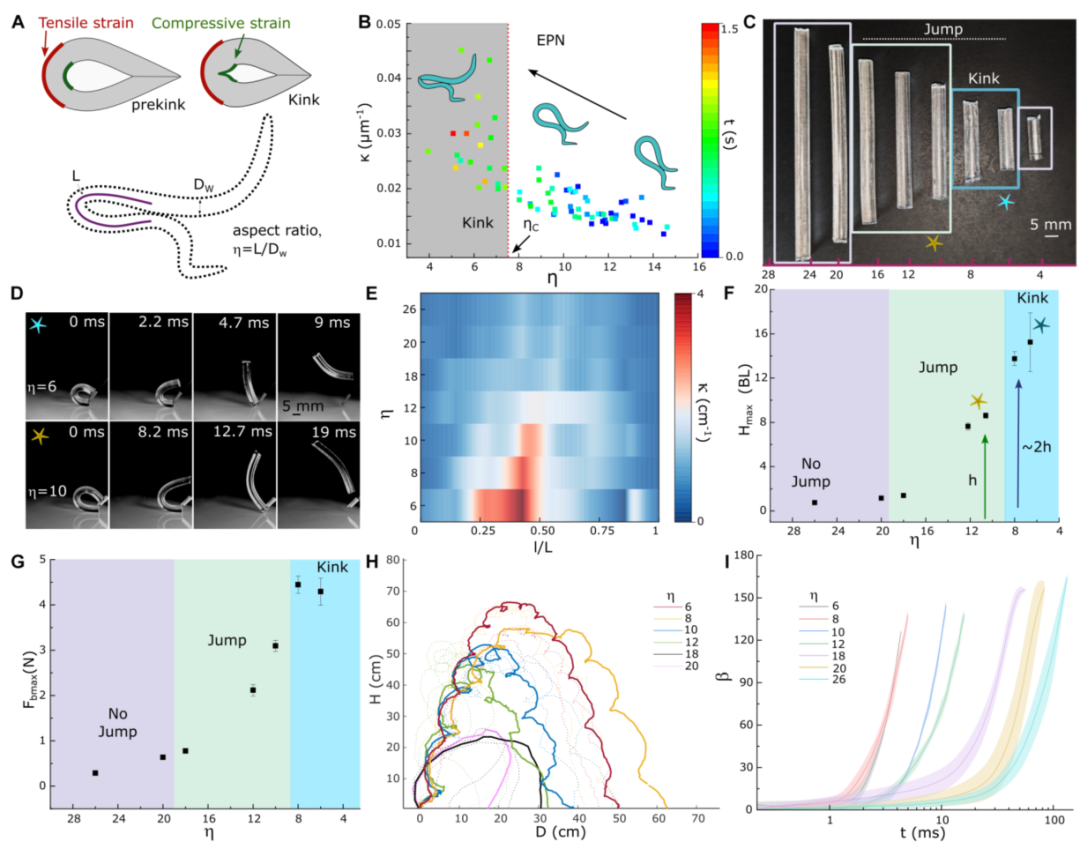
图6 纵横比在EPNs和SoftJM跳跃性能中的作用。(a)EPN的几何配置和从闭环到扭结形成的过渡,其中纵横比定义为η = 𝐿/𝐷w.。在弯曲或压缩应力期间,EPN的外表面承受拉伸应变,而内表面承受压缩应变。在临界弯曲度时,内表面会出现一个明显的扭结。(B)从环到纽结的EPN几何过渡,代表曲率κ和η之间的关系(n = 8个个体)。(C)对于1.2 MPa的模量,在不同η下的SoftJM 3,其中紫色方框显示在极低(η ≤ 4)和高(η ≥ 20)纵横比下没有跳跃。(D)soft JM 3的η = 6和η = 10的循环跳跃和打开。(E)在SoftJM 3弯曲和闭环形成期间,κ随η随标准化长度(l/L)的变化。(f)在不同η (n = 3,P < 0.0001)下,每bl soft JM 3的最大跳跃高度(𝐻max)。(G) 𝐹bmax形成一个闭环,η为SoftJM 3 (n = 3,P < 0.0001)。(H)不同η的SoftJM 3轨迹。(I)阴影误差曲线显示了SoftJM 3 (n = 3)的不同η的开度角(β)与时间的函数关系,阴影区域代表SD。
图6F和I进一步证实:长径比从12降到6(跨过临界值约7),跳跃高度从约5倍体长提升到约16倍体长,弹开速度也显著加快。换句话说,线虫主动压缩身体长径比,目的就是“越过”扭结的触发阈值,进入一种新的储能模式。
🚀刚柔并济的软体跳动机器人
基于这些机理,他们设计出SoftJM 4——碳纤维骨架+硅胶软体(图7A)。碳纤维模量约134 GPa,硅胶软体模量约几MPa,刚柔组合让结构既有储能所需的刚度,又有与人交互的安全性。
图7B-F展示了碳纤维数量对性能的影响:8根碳纤维(CF8)的跳跃速度达到约13米/秒,高度约3米(约25倍体长),功率输出20瓦/公斤,是1根碳纤维(CF1)的20倍。
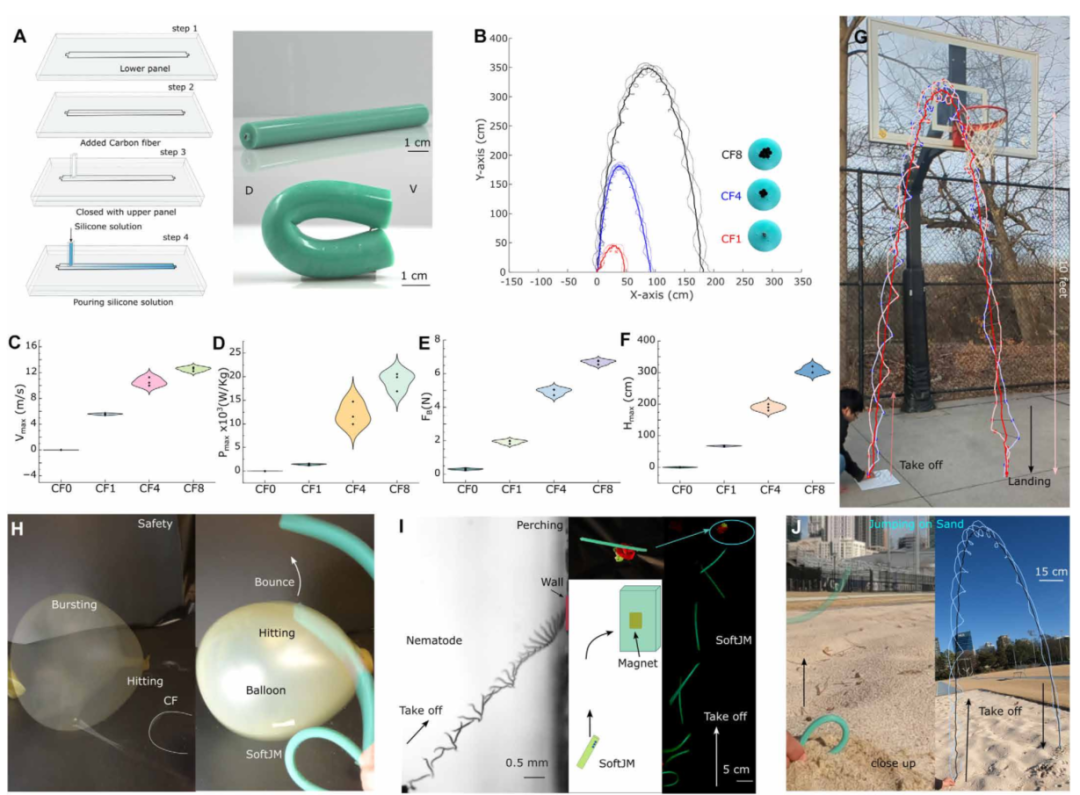
图7 SoftJM 4的多功能应用和安全演示。(A)soft JM 4的制造步骤,显示了CF-incorporated SoftJM,其中η = 24(详情见表S2)。(B)具有1、4和8 CFs的SoftJM的横截面视图,以及分别用红色、蓝色和黑色表示的CF1、CF4和CF8的SoftJM 4的完整轨迹,突出显示的线显示质心轨迹。(c到F) Violin图包括具有不同数量CFs的SoftJM 4的𝑉max的中心数据点、SoftJM 4在起飞期间产生的𝑃max、SoftJM 4的𝐹bmax以及各种软跳跃模型的𝐻max比较,其中n = 3,在CF1、CF4和CF8之间,P < 0.0001。(G) SoftJM 4 (CF8)跳过篮球框的高度(3.05米)。(H)安全特性表现为碳纤维迅速张开并使气球爆裂,而SoftJM 4在气球上弹跳而没有损坏。(一)EPN对寄主的伏击觅食行为。类似地,磁性软膜4表现出对磁性基底的吸引力。(J) SoftJM 4可以在沙子等颗粒状基质上跳跃。
他们还验证了几个实用性功能:图7G跳过一个3.05米高的篮球框(约25倍体长),图7H显示碳纤维骨架可以戳破气球,但被硅胶包裹后撞击气球只会弹开,不会伤人。图7J展示了沙地跳跃能力——对软体机器人来说,在松软介质上起跳比硬地面更难,但SoftJM 4依靠足够的功率输出做到了。
03 创新点
①把“扭结”从失效变成驱动开关
过去,弹性结构的扭结通常被视为失效。这项研究第一次揭示了扭结如何作为一种可逆储能机制被生物系统主动利用,并给出了完整的力学模型和仿生验证。这为软体机器人提供了一种新的驱动范式:不追求“完美”的弹性变形,而是把非线性不稳定性变成可控的能量释放开关。
②液体门控:微秒级释能的物理锁扣
线虫用微米级水膜作为“锁扣”,70微秒内破裂释放能量。这种液体门控机制在生物系统中可能比我们想象的更普遍(比如真菌孢子释放),但在软体机器人设计中还很罕见。这项工作提供了一个可行的工程实现思路:用表面张力做可控“锁”,替代复杂的机械门闩。
③长径比作为主动调节参数
线虫通过改变身体几何(长径比从20降到7以下)来触发扭结、提升储能效率。这种主动调节几何参数来控制力学失稳的思路,对软体机器人的设计有直接启发——很多软体结构的设计往往固定几何,但未来或许可以把“形状变化”作为控制的一部分。
④刚柔并济的软体跳动机器人
SoftJM 4在保持软体安全性的同时,实现了3米跳跃高度(25倍体长),比大多数软体跳跃机器人的性能高出一个量级。关键是,它不是单纯堆砌刚度和动力,而是借鉴了线虫的“力饱和”储能策略——用有限的驱动力储存尽可能多的弹性能。
⑤系统性的跨尺度验证
从微米级线虫(生物实验)到毫米-厘米级软体模型(物理实验),再到数值模拟(Cosserat rod),三个尺度相互验证,机制清晰。这种跨尺度的研究方法对仿生机器人领域有参考价值:如果只做生物观测,结论停留在“现象描述”;如果只做工程模型,可能抓不住关键的物理机制。
04 总结与展望
从一根弯折的吸管,到一根会跳跃的线虫,再到一只能跳3米高的软体机器人——这条路径的每一步,都是对“不稳定性”的重新理解。传统的控制思维追求稳定,但这项研究提示我们:不稳定性可以被设计,可以被调控,也可以被用来存储和释放能量。
从工程角度看,这个工作的价值不只是提供了一个跳得高的软体机器人。它揭示了三个可推广的设计原则:力饱和储能(扭结让力不再随弯曲增长)、液体门控(表面张力做可控锁)、几何调节触发(改变长径比来开关失稳)。这些原则有可能用在更广泛的软体机器人设计中,比如抓取、推进、跳跃、甚至自修复结构。
从生物学角度看,这项工作也为“生物如何突破肌肉力极限”提供了一个具体案例。肌肉的力密度是有限的,但通过合理的几何和力学设计,生物可以像线虫这样把力量“放大”几个数量级。未来或许会发现更多类似机制——不只是在跳跃中,也可能在咬合、弹射、冲击等场景中。
未来研究将聚焦于以下几个方向:
🔸多扭结耦合与序列驱动:目前只有单扭结,如果能实现多个扭结的时序释放,可能实现多级弹射或更复杂的运动模式。线虫本身会不会用这种策略,值得进一步观察。
🔸液体门控的主动控制:目前水膜破裂是被动的(弯曲力超过毛细力)。如果能在表面引入可控湿润性,或者用电场/温度控制液体桥,就可能实现主动“预紧-释放”的精确时序。
🔸小型化与集成化:现在的SoftJM 4尺度在厘米级,碳纤维骨架是手工嵌入的。如果能用MEMS工艺把这种刚柔结构做到毫米级,可能用在微型机器人或医疗设备中。
🔸更多地形适应性:沙地已经验证了,但如果是更复杂的非平整表面(如粗糙岩石、植被),扭结驱动的跳跃是否仍然有效?需要更多实验。
🔸与感知和控制闭环结合:目前的跳跃是“开环”的——给定初始几何,弹开就结束了。如果加入传感器和主动控制,或许可以实现落地后自主复位、多次跳跃、甚至调整方向。
你设计的软体机器人,会不会也“打一个结”来跳得更高?在仿生机器人中,你更倾向于“避免”不稳定性,还是“利用”不稳定性?欢迎在评论区分享你的看法。
声明:本文仅供学术交流,版权归原作者所有。如有错误或侵权,请联系更正或删除,欢迎留言探讨。